


Our research group investigates how to create and renew the stem cells that maintain the tissues under physiological conditions. These processes have great similarities with those used by the tumours for their perpetuation. Our studies focus on the hematopoietic system and in the process of leukaemia.
Moreover, we work very closely with the Research Group for Molecular Mechanisms of Cancer and Stemness, directed by Dr. Lluís Espinosa in the transfer of our discoveries on haematopoietic cells to epithelial tissues such as gut and skin and vice versa. In addition, this partnership allows us to achieve a deep molecular understanding of the biological problems.
Our group was created more than a decade ago, and we have achieved scientific goals of the highest level in the field of Biomedical Sciences, as well as contributed to the training of new researchers, who are currently working on various National or International centres and Companies. In recent years, we have developed various experimental methods to address at all levels specific biomedical problems.

In fact, our experimental approach ranges from more biochemical research to the analysis of animal models that reproduce the pathologies of interest and allow us to confirm the functional relevance of our findings. The ultimate goal will be to study possible therapeutic applications through the analysis of patient samples. In this sense we have devoted special efforts to understand the regulation of haematopoietic stem cells, as a tool to understand the mechanisms that regulate leukemia.
Molecules that control both tumor initiation and progression, and tissue/stem cell homeostasis include those of the Notch, Wnt and NFκB signalling payhways. Studying these pathways in the hematopoietic, intetinal, and skin tissues has also led to the identification of several interactions between them, which proved to be crucial for tissue homeostasis. We are currently working on several projects that deal with different functional aspects of normal haematopoietic stem cell regulation as well as leukemia initiating cells.

Our group was pioneer in revealing that specific elements of the Notch and Wnt/b-catenin pathways are required for the generation of hematopoietic stem cells during embryogenesis. Our current studies are focused on understanding the signals that the embryo uses to form these self-renewing cells that maintain the hematopoietic system throughout the life of the organism.
Currently we use mutant mouse strains and several fluorescent reporter proteins, combined with cutting edge imaging technologies, cell culture and “in mouse” hematopoietic transplantion assays, to reveal how this process occurs. Collaborations with other groups (e.g. Pr. Pablo Menéndez, Josep Carreras Institute) have also facilitated the validation of our research and results in models of human embryonic stem cells.
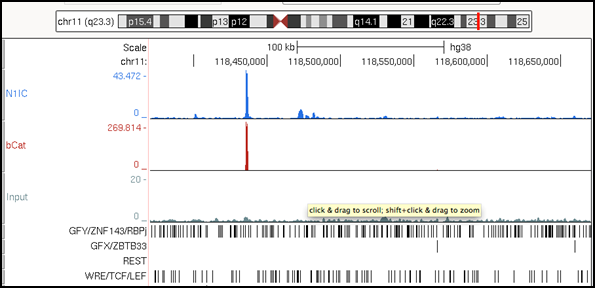
We study the signals that regulate the generation and maintenance of normal and leukemic cells, as well as leukaemic stem cells (LSCS). With this aim we have developed different in vitro and in vivo experimental models. In addition, we are now using several next-generation techniques to identify the gene expression patterns (RNA-seq), epigenetic patterns ( Chip-Seq ), and specific multi-protein complexes (mass spectrometry) that define the populations of interest at the molecular level. These studies may reveal novel therapeutic targets for T-ALL.
Leukaemia in neonates is more aggressive than that occurring in older children, which may be due to its pre-natal origin and the characteristics of the cells that undergone transformation. Our research is focused on determining the leukemic potential of hematopoietic cells at different stages/ages. Our research studies have focused on models of leukemia with MLL rearrangements but also in T-ALL leukemia induced by active Notch1. Our collaboration with the group of Dr. Mireia Camos in the children's Hospital Sant Joan de Déu is crucial in order to transfer our knowledge to the clinical practice.

Coordinator:
Anna Bigas(ELIMINAR)
Tel:
93 316 04 40
Fax:
93 316 04 10
Dr. Aiguader, 88
08003 Barcelona
© Institut Hospital del Mar
d'Investigacions MèdiquesLegal Notice and Privacy Policy | Cookie Policy | Site Index | Accessibility | Find Us | Contact
